

Het brein als spanningsmodulator: Ritmische veldprojectie als grondslag van bewustzijn, geheugen en ziekte
Dit artikel presenteert een alternatieve benadering van het brein, niet als opslagstructuur, maar als spanningsmodulerend orgaan dat ritmische veldprojecties organiseert. Herinnering, cognitie en pathologie worden hierin beschreven als dynamische ritmes van veldspanningen. Burn-out, dementie en lichamelijke aandoeningen worden herleid tot verstarde of verstoorde spanningspatronen. Gezondheid wordt gedefinieerd als het vermogen tot vrije ritmewisseling en veldcoherentie.
WETENSCHAPPELIJK
Paul Hager
7/9/202518 min lezen
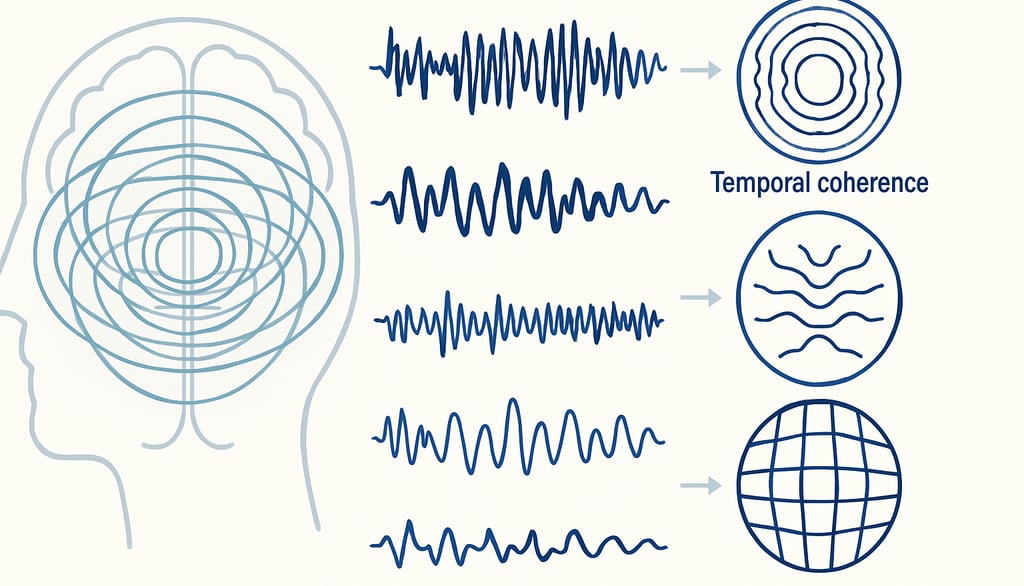
Inleiding — Het brein als spanningsmodulator: Weg van het opslagmodel
Binnen de hedendaagse neurowetenschap domineert nog steeds het idee van het brein als opslag- en verwerkingscentrum. Vanuit dit model wordt aangenomen dat het brein informatie opslaat in specifieke netwerken en neuronale verbindingen, waarna deze gegevens later kunnen worden opgeroepen in de vorm van herinneringen, gedachten of handelingen (Squire, 2004). De hersenen worden zo voorgesteld als een biologische computer, waarin data wordt verwerkt, opgeslagen en teruggelezen.
Toch ontstaan er de laatste decennia toenemende aanwijzingen dat deze metafoor tekortschiet. Diverse studies tonen aan dat herinneringen niet statisch liggen opgeslagen, maar dynamisch worden gereconstrueerd, afhankelijk van de actuele toestand van het breinnetwerk (Dudai, 2012; Schacter, 2012). De zogeheten 'reconsolidatie' van herinneringen wijst op een voortdurend herordenen en bijstellen van geheugeninhouden, wat moeilijk te verenigen is met een lineair opslagmodel.
Daarnaast groeit het inzicht dat hersenactiviteit primair georganiseerd wordt door ritmische en oscillatoire processen. Hersengolven, variërend van delta- tot gammafrequenties, coördineren grootschalige netwerken en zijn cruciaal voor geheugen, aandacht en bewustzijn (Buzsáki, 2006; Fries, 2015). Deze ritmes moduleren niet alleen neuronale communicatie, maar vormen in toenemende mate de kern van de informatieverwerking zelf.
Vanuit dit perspectief wordt het brein zichtbaar als een spanningsmodulerend orgaan: een systeem dat ritmes genereert, spanning in de tijd verdeelt en daarmee ervaring structureert. In plaats van gegevensopslag en -ophaling, functioneert het brein als een verdeler van veldspanningen, waarbij ervaring ontstaat uit de dynamische ordening van ritmes over tijd.
Deze benadering sluit aan bij het bredere theoretische kader waarin bewustzijn en materie worden opgevat als patronen van veldcoherentie, zoals onder meer beschreven in de impliciete orde van David Bohm (1980) en in modellen van kwantum-resonantie bij Penrose en Hameroff (2011). Vanuit dit spanningsperspectief worden hersenfuncties niet langer herleid tot een intern opslagmechanisme, maar tot een ritmische projectie van veldstructuren, waarbij het brein fungeert als modulatie-interface.
Dit artikel werkt dit veldperspectief op het brein systematisch uit. Vanuit neurowetenschappelijke inzichten over ritmes en plasticiteit wordt beschreven hoe het brein ervaring mogelijk maakt door de differentiatie van spanning over tijd. Hierbij wordt onderzocht hoe geheugen, bewustzijn, emotie en ziekte kunnen worden begrepen als verschillende stadia van ritmische veldinteractie — en hoe verstoringen in deze ritmes kunnen leiden tot psychische en lichamelijke klachten, waaronder burn-out en dementie.
Door het brein te benaderen als spanningsmodulator, ontstaat een nieuw raamwerk waarin het begrip van bewustzijn, gezondheid en ziekte radicaal verschuift: van opslag en defect, naar ritme en veldcoherentie.
Dit artikel bouwt voort op het Noöhedral Model van Hager (2025), waarin bewustzijn wordt beschreven als een fundamenteel veld (Ψ) dat opereert in een Hilbert-ruimte, met dynamische parameters zoals spanningsgradiënten (∇Φ) en coherentie-index (λ). Dit kader biedt een wiskundige basis voor het begrijpen van hersenfuncties als ritmische modulaties van veldspanningen.
Hoofdstuk 1 — Kader: Wat betekenen veldresonantie en spanningscirculatie?
Om de concepten in dit artikel toegankelijker te maken, leggen we hier twee kernbegrippen uit in eenvoudige termen:
Veldresonantie: Stel je voor dat het brein werkt als een orkest. Gedachten, emoties en herinneringen zijn als muziekinstrumenten die golven van activiteit produceren. Veldresonantie is het proces waarbij deze golven op elkaar afstemmen, zoals wanneer een orkest harmonieus samenspeelt. In dit artikel beschrijven we dit als de dynamische ordening van spanningen in het brein, die ervaringen zoals bewustzijn en geheugen mogelijk maakt.
Spanningscirculatie: Zie het brein als een rivier waarin water (energie) stroomt. Spanningscirculatie verwijst naar de manier waarop deze energie door het brein wordt verdeeld en georganiseerd in ritmische patronen, zoals golven in een rivier. Deze patronen sturen hoe we denken, voelen en handelen, en verstoringen hierin kunnen leiden tot problemen zoals burn-out of dementie.
Deze analogieën, geïnspireerd door het Noöhedral Model (Hager, 2025), helpen om de abstracte ideeën van dit artikel te begrijpen als een nieuwe manier om naar het brein te kijken: niet als een computer, maar als een dynamisch, ritmisch systeem.
Hoofdstuk 2 — Het brein als spanningsmodulator: Ritmische verdeling als functioneel principe
De kernfunctie van het brein wordt in de traditionele neurowetenschap vaak samengevat als informatieverwerking. Vanuit die benadering worden sensorische prikkels gecodeerd, getransporteerd, verwerkt en uiteindelijk omgezet in gedrag of herinnering. Maar deze lineaire kijk op hersenactiviteit verliest snel terrein.
Moderne neurofysiologische studies laten zien dat hersenactiviteit zelden lineair of sequentieel verloopt. Integendeel, de hersenen blijken te functioneren als een complex, zelforganiserend systeem waarin ritmes en oscillaties de fundamentele organiserende krachten zijn (Buzsáki, 2006; Varela et al., 2001). Deze ritmes vormen geen bijproduct van neuronale activiteit, maar liggen aan de basis van de coördinatie van grootschalige netwerken.
Hersengolven in verschillende frequenties — zoals delta (0,5–4 Hz), theta (4–8 Hz), alfa (8–12 Hz), bèta (12–30 Hz) en gamma (>30 Hz) — blijken direct betrokken bij processen zoals waakzaamheid, geheugen, motoriek, aandacht en bewustzijn (Fries, 2015; Buzsáki, 2006). Daarbij is niet alleen de frequentie van belang, maar ook de mate van synchronisatie tussen verschillende hersengebieden.
Het brein werkt hierin als spanningsmodulator: het verdeelt inkomende en interne spanning over tijd, waarbij specifieke hersenritmes fungeren als functionele vensters waarin informatie wordt geïntegreerd en beschikbaar wordt gesteld voor ervaring. In deze optiek is het brein geen opslagmedium, maar een dynamische verdelingsruimte waarin veldspanningen worden geordend tot herkenbare patronen.
Dit proces is direct zichtbaar in studies naar hersenplasticiteit, waarin blijkt dat de hersenen zich voortdurend herschikken in reactie op nieuwe ervaringen. Hersennetwerken reorganiseren zich op basis van ritmische stimulatie en veldcoherentie, niet door statische gegevensopslag (Kolb & Whishaw, 1998; Sporns, 2011).
Belangrijk is hierbij dat hersenactiviteit niet kan worden begrepen als lokale gebeurtenis. Neuroimaging laat zien dat cognitieve functies ontstaan vanuit grootschalige interacties tussen netwerken die zichzelf moduleren via oscillatoire koppeling (Deco et al., 2011). Deze netwerken vormen geen vaste circuits, maar tijdelijke samenklontering van velddynamiek die afhankelijk is van de actuele ritmische toestand.
Het brein maakt hiermee geen ‘beslissingen’ in de klassieke zin. Het vergelijkt, weegt en verdeelt spanning over ritmische cycli, waardoor ervaringen ontstaan als interferentiepatronen van verschillende veldritmes.
Vanuit dit veldperspectief kan het brein worden beschouwd als een dynamische veldmodulator:
Het herordent inkomende spanning via ritmische oscillaties.
Het genereert temporele structuur waarin ervaring mogelijk wordt.
Het fungeert als koppelpunt tussen externe zintuiglijke stromen en interne veldpatronen.
|
Deze benadering sluit nauw aan bij beschrijvingen van het brein als dissipatief systeem, waarin patronen van resonantie en zelforganisatie bepalend zijn voor bewustzijn en gedrag (Prigogine, 1997; Freeman, 2000).
Samengevat: de functie van het brein ligt niet in opslag, maar in het ritmisch moduleren van spanning over tijd. Wat wij ervaren als bewustzijn, geheugen of denken, is het resultaat van deze continue veldverdeling, waarin patronen van coherentie ontstaan en verdwijnen binnen de ritmische dynamiek van het brein.
Hoofdstuk 3 — Herinnering als veldherhaling: Geheugen als ritmische resonantie, geen opslag
Binnen het klassieke neuropsychologische model wordt geheugen doorgaans benaderd als een proces van opslag en reproductie. Herinneringen zouden als stabiele ‘geheugensporen’ (memory traces) worden vastgelegd in synaptische netwerken, waarna ze bij latere activatie weer toegankelijk worden gemaakt (Squire, 2004).
Hoewel het opslagmodel, gebaseerd op synaptische plasticiteit (Hebb, 1949), waardevolle inzichten biedt in lokale geheugenprocessen, interpreteert het Noöhedral Model deze als emergente eigenschappen van bredere velddynamica (Hager, 2025). Neurochemische signalering kan worden gezien als een manifestatie van spanningsmodulaties binnen Ψ(t). Ψ(t) vertegenwoordigt de dynamische evolutie van het bewustzijnsveld in de tijd, zoals beschreven in Hager, 2025
Deze benadering vindt haar wortels in het werk van Donald Hebb (1949), die het principe van synaptische plasticiteit introduceerde. Volgens dit model wordt een herinnering gevormd door het versterken van synaptische verbindingen tussen neuronen die gelijktijdig actief zijn geweest — het zogenaamde ‘cells that fire together, wire together’-principe.
Toch hebben tal van recente studies dit opslagmodel ter discussie gesteld. Onderzoek naar geheugenreconsolidatie toont aan dat herinneringen geen vaste entiteiten zijn, maar dynamische processen. Wanneer een herinnering wordt opgeroepen, wordt zij tijdelijk labiel, gevoelig voor wijziging en herschikking, alvorens opnieuw te worden ‘geconsolideerd’ in het netwerk (Dudai, 2012; Nader & Einarsson, 2010). Herinneringen blijken dus geen statische gegevens, maar veranderlijke patronen die bij elke oproeping opnieuw worden gevormd.
Vanuit een veldperspectief is dit verschijnsel goed verklaarbaar.
Herinnering ontstaat niet uit het ophalen van opgeslagen data, maar uit het herhalen van velddynamiek. Wanneer je je iets herinnert, wordt niet een ‘bestand’ opgehaald, maar wordt een specifiek spanningspatroon opnieuw aangeslagen in het brein, waardoor dezelfde veldcoherentie tijdelijk terugkeert.
Hersengolven en oscillaties spelen hierin een centrale rol. Studies tonen aan dat specifieke frequenties, zoals theta-ritmes (4–8 Hz), sterk geassocieerd zijn met geheugenprocessen, vooral in de hippocampus en de prefrontale cortex (Buzsáki, 2006; Nyhus & Curran, 2010). Deze ritmes moduleren de timing van synaptische activiteit en creëren vensters waarin geheugenintegratie plaatsvindt.
Herinnering is in dit model een ritmische resonantie:
Wanneer het brein in een bepaalde ritmische toestand komt, wordt een spanningspatroon geactiveerd dat eerder is ontstaan.
Dit patroon is geen exacte reproductie, maar een nieuwe afstemming op het onderliggende veld, waarin de eerdere ervaring als spanningsconfiguratie ligt opgeslagen.
Herinneringen zijn daardoor nooit identiek, maar variabel, afhankelijk van de huidige veldcoherentie.
Deze benadering sluit aan bij neurodynamische modellen van geheugen, waarin wordt benadrukt dat herinneringen ontstaan vanuit de hernieuwde synchronisatie van netwerken in plaats van vaste opslag (Jirsa & Kelso, 2000; Rabinovich et al., 2006). Het geheugenproces is hierin een dynamische herordening van veldactiviteit, niet een passieve reproductie van statische gegevens.
Het verklaart ook waarom herinneringen gevoelig zijn voor context, stemming en zintuiglijke omgeving. Het veld waarin de herinnering wordt aangeslagen, bepaalt hoe het patroon wordt gereconstrueerd. Dit fenomeen, bekend als ‘state-dependent memory’, wordt vaak beschreven in de cognitieve psychologie (Tulving & Thomson, 1973), maar krijgt binnen het veldperspectief een directe dynamische betekenis: veldtoestand bepaalt veldherhaling.
Kort samengevat:
Geheugen is geen opslagproces, maar een herhaling van ritmische veldstructuren.
Het brein fungeert hierbij als modulatie-instrument, niet als gegevensarchief.
Herinnering ontstaat telkens opnieuw, als tijdelijk resonantiepatroon binnen de ritmische velddynamiek van het brein.
Dit inzicht ondermijnt het klassieke geheugenmodel en opent de weg naar een fundamenteel ander begrip van herinnering — als veldresonantie die ontstaat wanneer het brein spanning herhaalt in tijd, gestuurd door ritmische coherentie.
Dit concept wordt ondersteund door Hagers Noöhedral Model, waarin geheugen wordt gemodelleerd als een dynamische herhaling van spanningsconfiguraties binnen Ψ(t), beschreven door de Lagrangian
(Hager, 2025).
Hoofdstuk 4 — Ritmische regimes in het brein: Denken, gevoel en gewoonte als veldritmes
Wanneer we het brein benaderen als spanningsmodulator, wordt duidelijk dat het niet werkt volgens één enkel ritme. Integendeel, de hersenen functioneren als een complex samenspel van verschillende ritmische regimes, die elk hun eigen frequentie, diepte en functionele rol hebben. In deze context kunnen de fenomenen die wij aanduiden als denken, gevoel en gewoonte worden opgevat als onderscheiden ritmische patronen binnen de neurodynamiek van het brein.
Neurofysiologisch onderzoek laat zien dat cognitieve processen, affectieve toestanden en gedragsgewoonten corresponderen met specifieke hersengolven en oscillatoire netwerken (Buzsáki, 2006; Siegel et al., 2012). Deze ritmes zijn niet alleen functioneel verschillend, maar interfereren ook voortdurend met elkaar, waardoor ervaring ontstaat als een dynamisch veld van overlappende spanningspatronen.
Denken is primair verbonden met snelle oscillaties, zoals bèta- (12–30 Hz) en gamma-activiteit (>30 Hz). Deze ritmes worden geassocieerd met gerichte aandacht, werkgeheugen, taalproductie en cognitieve controle (Fries, 2015). Denken manifesteert zich in het brein als snelle, fluctuerende patronen van spanning, die voortdurend schakelen tussen verschillende netwerken.
Gevoel daarentegen is gekoppeld aan trager stromende ritmes, met name theta (4–8 Hz) en delta (0,5–4 Hz), die nauw verbonden zijn met limbische structuren zoals de hippocampus en de amygdala (Pape et al., 2005). Emoties ontstaan binnen deze langzame oscillaties als diepe, diffuse spanningsvelden die niet primair cognitief zijn, maar lichamelijk en somatisch worden ervaren.
Gewoonte opereert grotendeels via zeer trage oscillaties en structurele netwerkstabiliteit, zoals zichtbaar in de zogenaamde default mode network (DMN) en salience network. Deze netwerken tonen basale rustactiviteit en onderliggende automatische patronen van interne spanningsverdeling (Raichle et al., 2001; Seeley et al., 2007). Gewoontes zijn in deze zin laagfrequente veldcirculaties die zichzelf in stand houden, onafhankelijk van bewuste cognitieve processen.
Binnen dit spanningsmodel ontstaan denken, gevoel en gewoonte niet als afzonderlijke functies, maar als parallelle ritmische regimes:
Denken als snelle, fluctuerende ritmiek.
Gevoel als langzame, diepe oscillatie.
Gewoonte als uiterst trage veldcirculatie met hoge stabiliteit.
Deze regimes interfereren voortdurend met elkaar en kunnen elkaar versterken of onderdrukken. Zo kan een emotionele toestand de snelheid van denken vertragen of versnellen, terwijl ingesleten gewoontes het bereik van cognitieve flexibiliteit beperken. Deze interferentie verklaart waarom gedrag vaak niet rationeel of consistent is, maar fluctueert afhankelijk van de dominante ritmische toestand van het brein.
Neurodynamische modellen beschrijven dergelijke interacties als 'metastabiliteit', waarbij hersennetwerken voortdurend schakelen tussen verschillende coördinatietoestanden zonder in een vaste toestand te bevriezen (Kelso, 1995; Tognoli & Kelso, 2014). Vanuit een veldperspectief is dit herkenbaar als het samenspel van veldritmes die samen het ervaringscontinuüm vormen.
Samengevat:
Het brein kent geen hiërarchische functieverdeling, maar dynamische veldritmes.
Denken, voelen en gewoonte zijn te begrijpen als onderscheiden spanningsfrequenties binnen het brein.
Bewustzijn ontstaat uit de interferentie van deze ritmische regimes, waarin het brein als modulator de spanningspatronen in tijd ordent.
Dit inzicht biedt een alternatief voor klassieke psychologische modellen en plaatst cognitieve en emotionele functies in een gemeenschappelijk dynamisch spanningsraamwerk, waarin het onderscheid tussen ‘mentaal’ en ‘somatisch’ vervaagt.
Hoofdstuk 5 — Vastlopen van ritmes: Burn-out als ritmische fixatie en verlies van flexibiliteit
Binnen het spanningsmodel van het brein wordt duidelijk dat gezondheid niet primair samenhangt met de sterkte van individuele ritmes, maar met het vermogen tot ritmische flexibiliteit. Gezond functioneren vraagt een voortdurende afwisseling en dynamiek tussen de verschillende veldritmes van denken, voelen en gewoonte. Wanneer deze flexibiliteit verloren gaat en het brein langdurig in één dominant ritme blijft hangen, ontstaat een toestand van systemische verstarring, waarbij zowel de cognitieve als somatische velden worden belast.
Een typisch voorbeeld van een dergelijke ritmische fixatie is burn-out. Burn-out wordt traditioneel gezien als het eindstadium van chronische stress, gekenmerkt door uitputting, verminderde prestaties en affectieve vervlakking (Maslach et al., 2001). In neurologische termen wordt burn-out vaak in verband gebracht met hyperactivatie van de hypothalamus-hypofyse-bijnieras (HPA-as) en verhoogde activiteit in stressgerelateerde hersengebieden zoals de amygdala (Pruessner et al., 1999).
Vanuit een veldperspectief wordt burn-out echter niet primair gezien als gevolg van externe stressoren, maar als ritmische fixatie. In deze toestand blijft het brein langdurig gevangen in hoge frequenties, zoals overmatige bèta- en gamma-activiteit, wat samenhangt met voortdurende alertheid, controle en prestatiegerichtheid (Buzsáki, 2006). Tegelijkertijd raken trage ritmes, die normaliter voor herstel en integratie zorgen (zoals theta en delta), structureel onderdrukt.
Neuroimagingstudies bij burn-outpatiënten tonen daadwerkelijk verminderde activiteit in netwerken die met herstel en interne afstemming te maken hebben, zoals het default mode network en frontale controlecircuits (van der Werff et al., 2013). Het brein verliest daarmee zijn capaciteit om te schakelen tussen actieritmes en rustregimes. De hele neurodynamiek van het individu wordt gereduceerd tot een smalle frequentieband die het lichaam en de psyche voortdurend in een toestand van overbelasting houdt.
In termen van veldcoherentie betekent dit dat burn-out ontstaat wanneer een spanningscirculatie zó dominant wordt dat ze zichzelf in stand blijft houden, zonder ruimte te laten voor andere veldstromen. Dit is vergelijkbaar met dynamische verstarringsprocessen in zelforganiserende systemen, waar bepaalde oscillaties vastlopen in stationaire regimes (Prigogine, 1997).
Belangrijk is dat dit mechanisme niet alleen cognitieve klachten veroorzaakt, maar ook leidt tot fysieke manifestaties zoals slapeloosheid, spierpijn, hormonale disbalans en verminderde immuniteit. Het hele lichaam raakt afgestemd op het dominante, vastgelopen ritme, waardoor zowel psychische als somatische klachten ontstaan.
Herstel van burn-out is in dit licht geen kwestie van ‘rust nemen’ in klassieke zin, maar vereist het actief doorbreken van de vastgelopen ritmiek. Methoden die nieuwe ritmes in het systeem brengen, zoals gecontroleerde ademhaling, lichaamsgerichte therapieën, ritmische muziek of ritueel bewegingswerk, blijken in toenemende mate effectief (Santarnecchi et al., 2021). Zij herstellen niet simpelweg ‘balans’, maar forceren het brein om opnieuw te schakelen tussen verschillende spanningsfrequenties, waardoor veldcoherentie geleidelijk wordt herwonnen.
Samengevat:
Burn-out is een toestand van ritmische fixatie, waarbij het brein gevangen blijft in een hoge spanningsfrequentie.
Deze fixatie leidt tot het verlies van ritmische flexibiliteit en veroorzaakt zowel psychische als lichamelijke klachten.
Herstel vereist herintroductie van variatie in het veld, waardoor het brein weer leert schakelen tussen verschillende ritmes.
In dit perspectief wordt burn-out niet langer gezien als uitputting of falen, maar als een verstoring in de ritmische veldmodulatie van het brein, met duidelijke aanknopingspunten voor ritmisch-herstellende benaderingen.
Hoofdstuk 6 — Dementie en het verval van ritmische ordening: Tijdverlies en veldbehoud
Dementie wordt in de neurologie doorgaans gedefinieerd als een progressief verlies van cognitieve functies, met als kernkenmerk het verdwijnen van geheugen, taalvaardigheid en oriëntatievermogen (World Health Organization, 2020). In het dominante neurobiologische model wordt deze achteruitgang vooral toegeschreven aan structurele schade, zoals atrofie van de hippocampus, ophoping van amyloïde plaques en neurofibrillaire tangles (Jack et al., 2018).
Toch blijkt uit neurodynamisch onderzoek dat de kern van dementie niet uitsluitend ligt in structurele achteruitgang, maar vooral in het verlies van ritmische ordening. Functionele MRI- en EEG-studies tonen aan dat de vroege stadia van dementie gepaard gaan met ontregeling van hersenritmes, nog voordat structurele schade volledig zichtbaar is (Babiloni et al., 2016). Met name trage oscillaties zoals alfa- en theta-golven, die cruciaal zijn voor geheugen en oriëntatie, blijken progressief verstoord te raken (Jeong, 2004).
Vanuit een veldperspectief wordt dementie niet primair gezien als geheugenverlies, maar als een verlies van temporele veldstructurering. Het brein verliest hierbij zijn vermogen om inkomende spanningsstromen in de tijd te ordenen. Dit betekent dat het onderscheid tussen verleden, heden en toekomst geleidelijk vervaagt, niet door verlies van ‘geheugensporen’, maar door het wegvallen van de ritmische modulatie die deze tijdservaring mogelijk maakt.
In neurologische termen is dit zichtbaar in het progressieve wegvallen van functionele connectiviteit tussen hersengebieden die betrokken zijn bij temporele integratie, zoals de hippocampus, prefrontale cortex en pariëtale gebieden (Seeley et al., 2009). Vanuit veldcoherentie bezien betekent dit dat het brein niet langer in staat is om spanning tot een samenhangend tijdsritme te vouwen. Wat overblijft, is een fragmentarische stroom van ervaringen zonder duidelijke ordening.
Toch wijst onderzoek ook op iets opvallends. Bij mensen met gevorderde dementie blijven vaak diepe emotionele reacties mogelijk, vooral op muziek, aanraking of geur (Cuddy et al., 2012). Deze waarneming suggereert dat, ondanks het verlies van cognitieve temporele structuren, het onderliggende veld van spanning en resonantie intact blijft. Dit komt overeen met het veldmodel, waarin het brein slechts fungeert als temporale interface, terwijl het veld zelf — de bron van spanning en resonantie — niet aan neurobiologische aftakeling onderhevig is.
Dementie kan binnen dit perspectief worden opgevat als een ontvlechting van breinritme en veldstructuur:
De hersenen verliezen hun vermogen tot temporele modulatie, waardoor herinnering, oriëntatie en planning vervagen.
Het veldpatroon zelf blijft echter resonant, wat verklaart waarom basale emoties, muzikale gevoeligheid en affectieve reacties intact kunnen blijven.
Dementie is daarmee geen ‘verlies van zelf’, maar een verschuiving van cognitieve veldprojectie naar directe, niet-tijdgebonden veldbeleving.
Dit inzicht heeft praktische implicaties. Benaderingen die gericht zijn op non-verbale, ritmische en zintuiglijke stimulatie blijken bijzonder effectief in het onderhouden van contact met mensen met dementie (van der Steen et al., 2018). Ze sluiten direct aan op de niet-tijdgebonden lagen van het veld, waar spanning nog altijd beweegt, ook als de breinritmes zijn vervaagd.
Samengevat:
Dementie is primair een verstoring van ritmische tijdsstructurering, niet louter geheugenverlies.
Ondanks cognitief verval blijft veldresonantie vaak intact, zichtbaar in basale affectieve en zintuiglijke reacties.
Dit wijst op een onderscheid tussen breinprojectie en veldcoherentie, waarin het ‘zelf’ niet volledig verdwijnt, maar verschuift van tijdgebonden ervaring naar directe veldresonantie.
Dementie laat zo zien dat bewustzijn niet exclusief geworteld is in breinstructuur, maar dat het brein een modulatie-instrument is voor een dieper, niet-lineair veld waarin ervaring blijft voortbestaan, ook buiten tijd.
Hoofdstuk 7 — Ziekte als veldstolling: Pathologie als vastgezette spanningscirculatie
Ziekte wordt in de geneeskunde traditioneel opgevat als een defect in het lichaam, veroorzaakt door genetische, microbiële of omgevingsfactoren. Symptomen worden hierbij gezien als directe uitingen van cellulaire schade of biochemische ontregeling. Binnen deze benadering staat het lichaam centraal als materieel systeem dat herstel vereist via externe interventie (Westerhof & Keyes, 2010).
Vanuit het spanningsmodel van het brein en lichaam ontstaat echter een radicaal ander perspectief. Hierin is ziekte niet louter het gevolg van biologische defecten, maar een manifestatie van vastgezette spanningscirculaties. Wat zich op celniveau toont als pathologie, begint op veldniveau als een langdurig verstard spanningspatroon dat zijn beweeglijkheid heeft verloren.
Dit principe sluit aan bij de theorie van dissipatieve structuren binnen de thermodynamica van Prigogine (1997), waarin systemen die hun flexibiliteit verliezen, stollen in starre patronen. In biologische systemen wordt dit proces zichtbaar in de vorm van chronische ontstekingen, fibrose en degeneratieve ziekten, waarin weefsels hun elasticiteit en herstelvermogen verliezen (Nathan & Ding, 2010).
In neurologische contexten wordt dit spanningsstollingsproces duidelijk zichtbaar bij chronische stress en trauma, waar langdurige hyperactivatie van stressnetwerken leidt tot structurele veranderingen in de hersenen, zoals volumeverlies in de hippocampus en functionele verstarring van limbische circuits (McEwen & Gianaros, 2010). De spanningspatronen van het brein worden hierbij zodanig rigide dat ze zich niet langer kunnen herschikken, wat de weg effent voor zowel psychische als somatische aandoeningen.
Binnen dit veldkader ontstaat ziekte op een gelaagd proces:
Aanvankelijk is er een verstoring in de ritmische veldcirculatie — bijvoorbeeld door herhaalde stress, trauma of langdurige spanning.
Wanneer deze verstoring zich blijft herhalen, stolt het veldpatroon geleidelijk, wat leidt tot verstarring in het neurofysiologische systeem.
Uiteindelijk manifesteert zich deze verstarring in het lichaam als zichtbare pathologie — van spierspanning tot auto-immuunziekten of metabole aandoeningen.
Wat wij ‘ziekte’ noemen, is dan niet een losstaand biologisch probleem, maar het eindstadium van een veldstollingsproces dat begon als verlies van bewegingsvrijheid in het spanningsveld.
Deze benadering wordt bevestigd in somatische studies waar emotionele spanningspatronen — zoals onopgeloste angst of woede — een directe rol blijken te spelen in het ontstaan en verloop van lichamelijke klachten (Kiecolt-Glaser et al., 2002). Het lichaam fungeert hierbij als laatste ‘laag’ waarin diep verankerde spanningen zichtbaar worden, wanneer het brein en zenuwstelsel deze niet langer kunnen afvoeren.
Belangrijk is dat deze vorm van ziekte ook omkeerbaar kan zijn, zolang het onderliggende veldpatroon wordt heropend. Interventies die gericht zijn op veldherstel — zoals ademwerk, lichaamsgerichte therapieën of klank- en bewegingstechnieken — blijken in toenemende mate effectief in het mobiliseren van vastgezette spanningspatronen, met zowel psychische als somatische verbeteringen tot gevolg (van der Kolk, 2014; Mehling et al., 2011).
Samengevat:
Ziekte is het resultaat van een langzame stolling van spanningscirculaties die hun flexibiliteit hebben verloren.
Wat als biologische pathologie zichtbaar wordt, begon als veldverstarring, vaak door langdurige spanning of onopgelost trauma.
Herstel vereist heropening van het spanningsveld, niet enkel symptoombestrijding op materieel niveau.
Deze benadering doorbreekt de scheiding tussen psychisch en lichamelijk en plaatst ziekte in een dynamisch veldkader waarin spanningscirculatie, bewegingsvrijheid en veldcoherentie centraal staan.
Hager (2025) stelt dat ziekte kan worden begrepen als een lage-coherentietoestand (lage λ), met spontane remissie als herstel van veldcoherentie, meetbaar via biophoton-emissies en EEG-synchronie.
Hoofdstuk 8 — Slotbeschouwing: Bewustzijn, gezondheid en ziekte als dynamische velddynamiek
In dit artikel is het brein niet benaderd als opslagplaats of als biologische computer, maar als een spanningsmodulerend orgaan dat ritmes organiseert en veldspanningen over tijd verdeelt. Vanuit deze invalshoek ontstaat een radicaal ander perspectief op geheugen, bewustzijn en ziekte.
De kern van deze benadering is dat het brein geen vaste gegevens opslaat, maar spanningspatronen voortdurend herschikt en projecteert in ritmische vormen van ervaring. Herinneringen blijken in dit licht geen opgeslagen feiten, maar tijdelijke herhalingen van eerdere spanningsdynamiek, waarbij de specifieke toestand van het veld telkens bepaalt hoe een herinnering zich aandient.
Dit ritmische model maakt ook duidelijk waarom ervaring gelaagd en fluctuerend is. Denken, voelen en gewoonte zijn geen afzonderlijke breinfuncties, maar ritmische regimes die elk hun eigen spanningsfrequenties volgen. Bewustzijn ontstaat als interferentiepatroon tussen deze ritmes, waarin het brein als modulator de ervaring in tijd ordent.
Cruciaal is dat gezondheid binnen dit model niet wordt gezien als afwezigheid van ziekte, maar als het vermogen van het brein en lichaam om vrij tussen ritmes te kunnen schakelen. Burn-out is hierin een voorbeeld van ritmische fixatie, waarbij het systeem gevangen blijft in een versnelde spanningscyclus. Dementie daarentegen laat zien wat er gebeurt als het brein zijn temporele ordeningsvermogen verliest, terwijl de onderliggende veldresonantie blijft bestaan.
Het meest fundamentele inzicht uit dit model is echter de beschouwing van ziekte als veldstolling. Wat in de geneeskunde wordt geclassificeerd als biologische pathologie, blijkt vanuit veldperspectief een proces van spanningsverstarring te zijn. Langdurige, onopgeloste spanningspatronen kunnen zich letterlijk verankeren in het lichaam, waar ze op den duur zichtbaar worden als weefselziekten of chronische ontstekingen.
Dit model laat zien dat het klassieke onderscheid tussen ‘geest’ en ‘lichaam’ kunstmatig is. Zowel psychische als lichamelijke fenomenen zijn uitdrukkingen van dezelfde onderliggende veldbeweging, waarin ritme en spanningscoherentie de kern vormen. Bewustzijn, gedrag en ziekte ontstaan niet door statische structuren, maar door dynamische spanningsprocessen die zich moduleren via het brein.
Vanuit deze benadering wordt duidelijk:
Bewustzijn is een veldfenomeen, tijdelijk geprojecteerd via breinritmes.
Gezondheid is de flexibiliteit om vrij tussen veldritmes te bewegen.
Ziekte is gestolde spanning, een verlies van veldbeweeglijkheid.
Dit spanningsperspectief opent daarmee een nieuwe benadering van menselijk functioneren, waarin het brein niet langer wordt gezien als een machine of harde schijf, maar als een ritmisch veldorgaan dat continu de balans bewaart tussen bewegen en verstarren, tussen projectie en loslaten, tussen ritme en stilte.
Critici kunnen stellen dat het opslagmodel beter geschikt is voor het verklaren van specifieke geheugenprocessen, zoals de rol van de hippocampus in episodisch geheugen (Squire, 2004). Dit model, gebaseerd op synaptische plasticiteit (Hebb, 1949), biedt waardevolle inzichten in hoe lokale neurale netwerken herinneringen coderen en ophalen. Daarnaast benadrukken neurowetenschappers de cruciale rol van neurochemische processen, zoals dopaminerge en cholinerge signalering, in cognitie en emotie. Het spanningsmodel, ondersteund door Hagers Noöhedral Model (2025), biedt echter een breder kader waarin deze lokale processen worden geïnterpreteerd als projecties van veldcoherentie. In dit perspectief zijn synaptische plasticiteit en neurochemische signalering emergente eigenschappen van dynamische spanningsmodulaties binnen het bewustzijnsveld Ψ(t). Toekomstig onderzoek kan deze benaderingen integreren door bijvoorbeeld te onderzoeken hoe hersenritmes, zoals theta- en gamma-oscillaties, neurochemische signalen moduleren in samenhang met veldcoherentie (Hager, 2025, Appendix K). Dit opent de deur naar een holistische neurowetenschap die lokale en globale mechanismen verenigt.
Een ander mogelijk tegenargument is de huidige beperkte empirische validatie van het veldmodel. Hoewel Hager (2025, Appendix I) experimentele protocollen voorstelt, zoals biophoton- en EEG-metingen, zijn deze nog in de conceptfase, wat verdere validatie vereist." Dit zou de zelfreflectie verder versterken.
In deze visie wordt gezondheid geen statische toestand, maar een kunst van veldbeweging.
Referenties
Buzsáki, G. (2006). Rhythms of the brain. Oxford University Press.
Dudai, Y. (2012). The restless engram: consolidations never end. Annual Review of Neuroscience, 35, 227–247. https://doi.org/10.1146/annurev-neuro-062111-150500
Fries, P. (2015). Rhythms for cognition: communication through coherence. Neuron, 88(1), 220–235. https://doi.org/10.1016/j.neuron.2015.09.034
Jeong, J. (2004). EEG dynamics in patients with Alzheimer's disease. Clinical Neurophysiology, 115(7), 1490–1505. https://doi.org/10.1016/j.clinph.2004.01.001
McEwen, B. S., & Gianaros, P. J. (2010). Central role of the brain in stress and adaptation: Links to socioeconomic status, health, and disease. Annals of the New York Academy of Sciences, 1186(1), 190–222. https://doi.org/10.1111/j.1749-6632.2009.05331.x
Nathan, C., & Ding, A. (2010). Nonresolving inflammation. Cell, 140(6), 871–882. https://doi.org/10.1016/j.cell.2010.02.029
Penrose, R., & Hameroff, S. (2011). Consciousness in the universe: Neuroscience, quantum space-time geometry and Orch OR theory. Journal of Cosmology, 14. (Open access)
Prigogine, I. (1997). The end of certainty: Time, chaos, and the new laws of nature. Free Press.
Sporns, O. (2011). Networks of the brain. MIT Press.
van der Kolk, B. A. (2014). The body keeps the score: Brain, mind, and body in the healing of trauma. Viking.
van der Steen, J. T., Smaling, H. J. A., van der Wouden, J. C., Bruinsma, M. S., Scholten, R. J. P. M., & Vink, A. C. (2018). Music‐based therapeutic interventions for people with dementia. Cochrane Database of Systematic Reviews, 7. https://doi.org/10.1002/14651858.CD003477.pub4
Hager, P. (2025). The Noöhedron: A Differential Field Theory of Consciousness and Reality. Zenodo. DOI: 10.5281/zenodo.15367135.
Hager, P. (2025). Emergent Spacetime from Field Coherence: Quantum-Tensional Geometry in the Ψ-metric Framework without Background Dynamics. Zenodo. DOI: 10.5281/zenodo.15526936.
Hager, P. (2025) The Noöhedral Resolution of Spontaneous Remission, Zenodo. DOI: 10.5281/zenodo.15188855
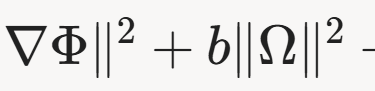
Over deze site
Verkenning van bewustzijn, materie en evolutie.
Paul Hager – Noöhedron
blijf op de hoogte
paul@noohedron.com
© 2025. All rights reserved.